Introducción a la microscopía de fluorescencia multifotónica
La microscopía de fluorescencia multifotónica es una poderosa herramienta de investigación que combina las técnicas ópticas avanzadas de la microscopía de escaneo láser con la excitación de fluorescencia multifotónica de larga longitud de onda a fin de capturar imágenes tridimensionales de alta resolución a partir de muestras marcadas con fluorocromos altamente específicos.

Figura 1. Configuración del microscopio de fluorescencia de excitación multifotónica
La metodología es particularmente útil para los biólogos celulares que se especializan en estudiar los procesos dinámicos en células y tejidos vivos sin infligir un daño significativo, a menudo letal, a la muestra. Si bien la microscopía de fluorescencia clásica de campo amplio puede proporcionar con frecuencia una resolución submicrónica de eventos bioquímicos en sistemas vivos, esta técnica tiene una sensibilidad y resolución espacial limitadas debido al ruido de fondo causado por la fluorescencia secundaria en áreas que se sitúan por encima y por debajo del plano focal.
La excitación en el caso de la microscopía multifotónica ocurre solo en el punto focal de un microscopio de difracción limitada, lo que brinda la capacidad de seccionar de forma óptica muestras biológicas gruesas para obtener una resolución tridimensional. Las secciones ópticas individuales son adquiridas mediante un escaneo de trama en el plano x-y, y una imagen 3D se compone del escaneo de la muestra en modo serial bajo posiciones secuenciales z. Puesto que la posición del punto focal puede ser determinada y controlada de forma precisa, la fluorescencia multifotónica es útil para analizar las regiones seleccionadas por debajo de la superficie de la muestra. La energía de excitación altamente localizada permite minimizar el fotoblanqueo de los fluorocromos que se adhieren a la muestra, como también reducir el fotodaño, lo que aumenta la viabilidad celular y la duración sucesiva de los experimentos centrados en investigar las propiedades de las células vivas. Adicionalmente, la aplicación de las longitudes de onda por excitación en el infrarrojo cercano favorece una penetración más profunda en los materiales biológicos y reducen el alto grado de dispersión de luz que se observa con las longitudes de onda más cortas. Estas ventajas permiten a los investigadores ejecutar experimentos en muestras gruesas de tejido vivo, como en cortes de cerebro o en el cerebro vivo de animales vivos in vivo, y desarrollar embriones que serían difíciles o imposibles de obtener imágenes con otras técnicas microscópicas.
En la Figura 1 se ilustra una configuración típica para experimentos de microscopía de fluorescencia multifotónica. El microscopio es un instrumento de estilo vertical diseñado para observar tejidos vivos de animales pequeños por experimentación in vivo. La parte posterior del cuerpo del microscopio es un sistema de láser pulsado con bloqueo de modo por titanio-zafiro: una de las fuentes preferidas para la excitación multifotónica debido a la alta intensidad máxima pero baja potencia promedio. Un sistema de detección basado en un fotomultiplicador de filtro se encuentra conectado a una posición lo suficientemente cercana al portaobjetivos del microscopio con el fin de detectar de manera efectiva la fluorescencia dispersa que ha sido capturada por el objetivo. Las imágenes digitales recopiladas por el microscopio son procesadas y analizadas por una estación de trabajo informática adjunta que puede ensamblar reconstrucciones tridimensionales a partir de secciones ópticas.
La microscopía de fluorescencia de campo amplio tradicional se ve cubierta de fluorescencia secundaria, generada lejos de la región focal, lo que contribuye al toque y la señal de alto ruido de fondo que a menudo oscurecen detalles importantes de la muestra. La microscopía confocal soluciona este problema en grado suficiente al rechazar la fluorescencia de fondo desenfocada mediante el uso de aberturas estenopeicas, cuyas secciones ópticas finas (menos de una micra) sin desenfoque se producen desde lo profundo de especímenes gruesos. La introducción de la microscopía de fluorescencia multifotónica ofrece una alternativa a la microscopía confocal a través de la excitación selectiva que se conjuga con una gama más amplia de opciones de detección. A diferencia de los microscopios confocales convencionales, el microscopio de la Figura 1 no requiere un estenopo cerca del detector para lograr la discriminación tridimensional, lo que aumenta radicalmente la eficiencia de las señales de fluorescencia emitidas. En el pasado, el alto costo y la complejidad de los sistemas de láser pulsado que se requerían para la excitación multifotónica limitaban el uso de la técnica. Hoy en día, los láseres y sistemas multifotónicos comerciales llave en mano han posicionado a la microscopía de fluorescencia multifotónica como el método de preferencia para muchas investigaciones.
Excitación bi- o trifotónica
Los principios básicos de la excitación multifotónica se describieron por primera vez hace más de 70 años cuando la Dr.a Maria Göppert-Mayer llevaba a cabo su investigación de tesis doctoral; sin embargo, la hipótesis no pudo confirmarse hasta la invención de los láseres de rubí pulsados, casi 30 años después. Con altas densidades de fotones, dos fotones pueden ser absorbidos simultáneamente (mediados por un estado virtual) al combinar sus energías para provocar la transición electrónica de un fluorocromo al estado excitado. Dado que la energía de un fotón es inversamente proporcional a su longitud de onda, los dos fotones deben presentar longitudes de onda de casi el doble con respecto a aquellas requeridas para la excitación de un solo fotón. Por ejemplo, si hay dos fotones que tienen una longitud de onda de 640 nanómetros (luz roja), estos pueden ser combinados para excitar un fluorocromo de absorción ultravioleta en la región (ultravioleta) de 320 nanómetros. El resultado será una emisión de fluorescencia secundaria de longitudes de onda más largas (azul o verde). Esta aplicación única significa que las longitudes de onda más largas, extendidas hasta la región infrarroja, pueden usarse convenientemente para excitar los cromóforos en un solo evento cuántico, los cuales emitirán posteriormente radiación secundaria en longitudes de onda más bajas.
La necesidad de dos fotones para cada evento de excitación requiere una constante de proporcionalidad que depende del cuadrado de la intensidad de excitación. A pesar de que los fotones no necesitan tener la misma longitud de onda para inducir la excitación multifotónica, la mayoría de los sistemas experimentales están diseñados con una sola fuente láser; por tanto, los dos fotones suelen ser miembros de una población definida que tiene una distribución de longitud de onda estrecha. A diferencia de la absorción que se produce con un solo fotón, la probabilidad de que un fluorocromo dado absorba de forma simultánea dos fotones es una función de la superposición espacial y temporal entre los fotones incidentes. Los cálculos basados en la suposición de que cada fluorocromo es expuesto a la misma sección transversal del láser indican que los fotones deben llegar en 10(-18) segundos (un attosegundo) entre sí. La escala temporal de esta superposición es consistente con el tiempo de vida de (10(-17) segundos o 0,01femtosegundo) del estado virtual intermedio.
En la fluorescencia multifotónica, se requieren altas densidades de fotones con el fin de asegurar un nivel suficiente para la excitación de los fluorocromos. De hecho, la concentración de fotones debe ser de casi un millón de veces la requerida para una cantidad equivalente de absorciones con un solo fotón. Esto se logra gracias a los láseres pulsados con bloqueo de modo de alta potencia, los cuales generan una cantidad significativa de energía durante los picos del impulso, pero poseen una potencia promedio lo suficientemente baja que evita daños en la muestra. Los pulsos breves, pero también intensos, emitidos por el láser aumentan la probabilidad de absorción bifotónica promedio en un fluorocromo específico bajo un nivel constante promedio de la potencia láser incidente. Reducir el nivel de potencia de excitación promedio, disminuye la cantidad de absorción a partir de un solo fotón, lo que también ocurre en la muestra durante la excitación. Los eventos de excitación monofotónica son los que conllevan en gran medida al calentamiento y a un porcentaje del fotodaño que ocurre en los experimentos de fluorescencia.
Las configuraciones típicas del láser pulsado emplean ciclos de trabajo cortos de alrededor 100 femtosegundos (10 e(-13) segundos) con un índice de repetición de 80 a 100 MHz destinado a experimentos de fluorescencia multifotónica. Este sistema favorece la adquisición exitosa de imágenes sin someter la muestra a una cantidad excesiva de calor ni fotodaño. La escala temporal para cada impulso, pese a ser denominado «ultracorta», sigue siendo de cuatro a cinco órdenes de magnitud más larga que el tiempo de reacción para la absorción bifotónica. La población de estados singlete en los cromóforos, excitados por un pulso de dos fotones, es idéntica a la obtenida durante la microscopía de fluorescencia confocal o de campo ancho convencional. Por consiguiente, la emisión de la fluorescencia secundaria después de la excitación bifotónica es indistinguible de la observada en los experimentos de un solo fotón. Un fluorocromo, como la rodamina, emitirá el mismo amplio intervalo de longitud de onda a partir de la fluorescencia secundaria indistintamente de un evento de excitación mono o bifotónica.
Diagrama de excitación multifotónica de Jablonski
Descubra cómo se producen los eventos de excitación de fluorescencia en la microscopía de uno, dos y tres fotones mediante el diagrama clásico de Jablonski.
La excitación de tres fotones o trifotónica es un evento de absorción óptica no lineal relacionada que puede ocurrir de manera similar a la excitación bifotónica. La diferencia es que tres fotones deben interactuar simultáneamente con el fluorocromo para provocar una transición al estado de singlete excitado. Un beneficio de la excitación trifotónica es que, para una absorción exitosa, se requiere solo una concentración de fotones diez veces superior a la absorción bifotónica, lo que hace que esta técnica sea interesante para algunos experimentos. La excitación trifotónica favorece la resolución del eje z en un grado aún mayor que la absorción de dos fotones. Esto se debe a una sección transversal más pequeña dedicada a la excitación del fluorocromo que resulta de la necesidad de una interacción simultánea entre tres fotones individuales. En la práctica, un láser que emite luz infrarroja con una distribución de longitud de onda centrada en 1050 nanómetros es capaz de excitar un fluorocromo que absorbe fotones en la región ultravioleta (alrededor de 350 nanómetros: un tercio de la longitud de onda de excitación). El mismo láser puede excitar simultáneamente otro fluorocromo a la mitad de la longitud de onda (525 nanómetros): una combinación útil en experimentos biológicos de doble marcado.
Al utilizar longitudes de onda más cortas (hasta 720 nanómetros) en el infrarrojo cercano, la fluorescencia de tres fotones puede extender la gama útil del procesamiento de imágenes de fluorescencia al ultravioleta profundo. Las longitudes de onda del láser en el intervalo de 900 a 700 nanómetros excitarán los fluorocromos que absorben en la región de 240 a 300 nanómetros, lo que es prácticamente inaccesible con los objetivos microscópicos convencionales. El vidrio que viene siendo usado en la fabricación de objetivos de fluorescencia presenta una transmisión muy baja para longitudes de onda inferiores a 300 nanómetros, pero la radiación láser infrarroja de longitud de onda más larga puede pasar fácilmente para producir una excitación trifotónica.
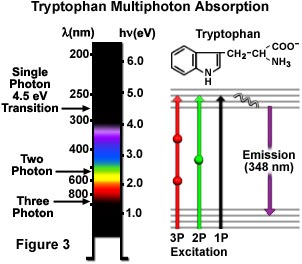
En la figura 3, se ilustran de forma esquemática las excitaciones de uno, dos y tres fotones del triptófano, un aminoácido aromático común. Una transición electrónica de un fotón de 4,5 electronvoltios excita el triptófano a 280 nanómetros mediante la posterior emisión de fluorescencia secundaria a 348 nanómetros en la región ultravioleta. La excitación generada por el mecanismo bifotónico se cumple con una luz de color amarillo verdosa centrada en 580 nanómetros, mientras que la excitación trifotónica ocurre cuando el aminoácido se ilumina con la radiación de 840 nanómetros en la región del infrarrojo cercano. Las transiciones se presentan en un diagrama de Jablonski (Figura 3), donde el estado virtual está representado por una esfera en el caso de la excitación bifotónica y, por dos esferas, en la trifotónica. El triptófano presenta una fluorescencia mucho más fuerte con un rendimiento cuántico superior en comparación con otros aminoácidos aromáticos; además, se halla sólo en pequeñas cantidades en la mayoría de las proteínas. Estas características convierten a la microscopía multifotónica en una excelente herramienta para las investigaciones que usan la autofluorescencia de los residuos del triptófano. Incluso otros fenómenos no lineales de orden superior son posibles, como la excitación cuatrifotónica, pero aún no han sido aplicados a la investigación biológica.
Microscopía de fluorescencia bifotónica
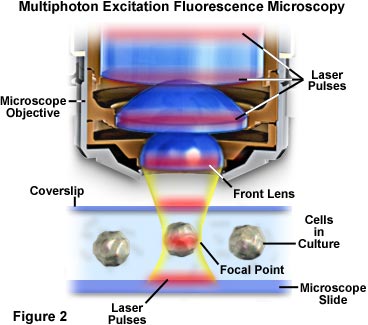
La localización de la excitación, en la región que circunda de inmediato al punto focal, en la microscopía multifotónica, ocurre porque es ahí donde la densidad de fotones es más alta. Esta ventaja surge del principio físico básico alusivo a que la absorción de dos fotones por un fluorocromo es una función del cuadrado de la intensidad de excitación. Cuando los fotones de una fuente de láser pulsado son enfocados por un objetivo de alta apertura numérica, estos se aglomeran más, lo que aumenta la probabilidad de que dos o más interactúen simultáneamente con un solo fluorocromo. La concentración de fotones en el punto focal del microscopio es tan crucial para la absorción multifotónica que ésta es la única región donde ocurre una excitación apreciable. El concepto se presenta en las Figuras 2 y 4, que ilustran la excitación multifotónica a nivel macroscópico y microscópico, respectivamente. La Figura 2 muestra una vista desproporcionada de un objetivo microscópico en posición, dedicado a capturar imágenes a partir de células cultivadas en un portaobjetos y cubreobjetos microscópicos. Los impulsos láser de luz roja atraviesan el eje longitudinal del objetivo; a continuación, se enfocan y concentran en la célula, en la parte central de la figura.
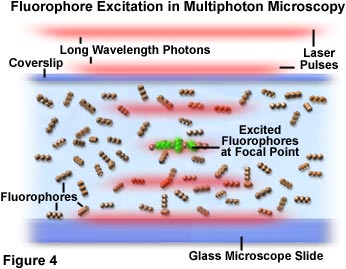
En la Figura 4, se demuestra la aglomeración de fotones y la interacción con los fluorocromos en el punto focal del microscopio. A medida que los impulsos láser de luz roja atraviesan la muestra que contiene fluorocromos (representados como un triplete lineal de esferas), la probabilidad de excitación aumenta a la vez que los impulsos alcanzan el punto focal del objetivo. Los fotones individuales se representan como un agregado segregado en líneas rojas difusas que definen los límites de los impulsos láser. En la Figura 4, un pequeño grupo de moléculas de fluorocromos, ubicadas en el centro de la región focal, han sido excitadas por la absorción simultánea de dos fotones y exhiben una fluorescencia secundaria verde. Hay una probabilidad muy baja de que los cromóforos al exterior del plano focal absorban dos fotones. Esto se debe a que la densidad de fotones no es lo suficientemente alta en esta región.
El fenómeno de la excitación bifotónica es posible no solo por la proximidad espacial de los fluorocromos en el punto focal del microscopio, sino también por la superposición temporal de los fotones contenidos en los impulsos láser secuenciales. Tal y como se ha mencionado anteriormente, la energía de excitación en la absorción de dos fotones ocurre en proporción al cuadrado de la intensidad del fotón producido por la fuente láser. La intensidad del rayo láser pulsado disminuye de acuerdo con el cuadrado de la distancia a partir del plano focal; por tanto, la probabilidad de excitación de un fluorocromo en cualquier lugar cerca de la región focal desciende a la cuarta potencia de la distancia del fluorocromo desde el plano focal. Las dimensiones del cono de iluminación láser pulsado se determinan por la apertura numérica objetiva. Por lo tanto, la disminución de la intensidad del haz lejos del punto focal es proporcional al cuadrado del diámetro del cono de la luz de excitación. A medida que el cono de iluminación se expande por encima y por debajo del punto focal, las probabilidades de excitación del fluorocromo disminuyen de acuerdo con la cuarta potencia del diámetro del cono. Por esta razón, la excitación del fluorocromo se limita a la región inmediata que rodea al punto focal, la cual representa solo una sección óptica muy delgada de la muestra completa.
Eventos de excitación regional
Examine los eventos que ocurren en la región focal del microscopio durante la excitación de la muestra mediante la iluminación láser visible e infrarroja cercana con longitud de onda larga.
La duración de los impulsos láser, que suele oscilar entre 100 femtosegundos y 1 picosegundo (10 e(-13) a 10 e(-12) segundo), es considerada ultracorta en términos macroscópicos. Sin embargo, en términos de escala temporal para los eventos de absorción de fotones (casi una milésima de femtosegundo), los impulsos ofrecen una duración bastante larga. Esto impide la saturación de fluorocromo y permite que las moléculas tengan el tiempo suficiente para volver al estado fundamental entre los impulsos, antes del empalme con la siguiente ronda de excitación. Los índices de repetición de impulsos oscilan entre 80 y 120 megahercios (MHz), lo que proporciona un alto pico de potencia instantáneo en la excitación, seguido de un tiempo de permanencia promedio de 10 nanosegundos. Dado que el tiempo de vida de la fluorescencia en un fluorocromo típico dura solo un par de nanosegundos, la población de moléculas excitadas dispone de mucho tiempo para relajarse entre los impulsos. El ciclo de trabajo relativamente corto del impulso (tiempo de duración del impulso dividido por el tiempo entre impulsos) limita la potencia láser promedio de entrada a un valor un poco mayor que aquel empleado habitualmente para la microscopía confocal de escaneo/barrido láser.
La limitación de la excitación bifotónica a la región cercana del plano focal proporciona una ventaja significativa de la microscopía multifotónica sobre la confocal. Con la microscopía confocal, la fluorescencia se excita en toda la muestra; pero, la fluorescencia secundaria recolectada por el detector se limita al plano focal del objetivo por el estenopo confocal. Esto permite reducir la cantidad de ruido de fondo o fluorescencia a partir de otros planos focales que agregan ruido de fondo a los datos. En cambio, la microscopía multifotónica genera excitación de fluorescencia (y, posteriormente, emisión de fluorescencia) solo en el plano focal, lo que elimina tanto la señal de fondo como la necesidad de un estenopo confocal. La gran diferencia entre los modos de excitación de microscopía confocal y multifotónica se ilustra en la Figura 5, que revisa los perfiles de fotoblanqueo con cada técnica.
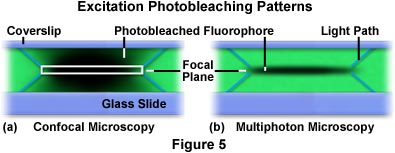
En la Figura 5 se presentan los patrones de fotoblanqueo x-z, producidos a partir del análisis repetido de un solo plano x-y en una película polimérica Formvar teñida con el fluorocromo rodamina (tinción verde). A la izquierda [Figura 5(a)], se halla perfil generado al escanear la película teñida con un microscopio confocal. El rectángulo blanco en el centro del escaneo representa el plano focal que pasa a través del estenopo y se ve reflejado por el detector. Las líneas azules diagonales que se proyectan desde las esquinas superior e inferior del rectángulo representan la trayectoria de la luz aplica por el haz luminoso de excitación a través de la película. Mientras la trama del haz escanea la película, la tinción fluorescente se excita y emite una fluorescencia secundaria. Al final, ocurre el fotoblanqueo, que está representado por las áreas oscuras en la región focal. En la película escaneada por el microscopio confocal (Figura 5[a]), la excitación integrada es casi igual a través de toda la trayectoria de excitación, tanto por encima como por debajo del plano focal. Por el contrario, el perfil de excitación del escaneo repetitivo x-z —generado por el microscopio multifotónico— limita la excitación y el fotoblanqueo al plano focal [Figura 5(b)]. De forma similar al caso de la Figura 5(a), las líneas diagonales azules que se producen a partir del plano focal trazan la trayectoria aplicada por la luz de excitación hasta llegar al plano focal.
La excitación localizada, facilitada por la microscopía multifotónica, crea enormes beneficios. Quizá el más significativo es el alto grado de resolución tridimensional que puede lograrse con esta técnica, la cual es idéntica a la que podría obtenerse con un microscopio confocal idealizado. Asimismo, la falta de absorción de los fluorocromos puestos en la parte externa del plano focal permite que más luz de excitación penetre a través de la muestra y alcance el plano de enfoque. El resultado es una capacidad altamente superior proporcionada por el haz enfocado que penetra profundamente dentro de la muestra, la cual puede alcanzar con frecuencia de dos a tres veces más la observada con la microscopía confocal.
Tal y como se abordó previamente, la probabilidad de absorción multifotónica fuera de la región focal disminuye según la cuarta potencia de la distancia a lo largo del eje óptico (dirección z). Cuando una distribución uniforme de fluorocromos se somete a una excitación multifotónica mediante un objetivo de alta apertura numérica (1.4), cerca del 80 % de la absorción se generará en un espacio estrechamente definido denominado volumen focal. Las dimensiones de este volumen dependen de la apertura numérica del objetivo; sin embargo, en el caso de un objetivo típico de fluorescencia con gran apertura en las longitudes de onda del infrarrojo cercano, esta área se definirá por un elipsoide con dimensión lateral de 0,3 micras de diámetro y una longitud axial de 1 micra.
Patrones de blanqueo por excitación
Compare patrones de fotoblanqueo inducidos por la excitación que se producen cerca de la región focal en sistemas de microscopía confocal y multifotónica.
La importante reducción del fotoblanqueo (y fotodaño asociado a células y tejidos), ilustrada en la Figura 5(b), que proporciona la microscopía multifotónica es superior a la que facilita la microscopía confocal. El fotoblanqueo y el fotodaño son dos de las limitaciones más importantes de la microscopía de fluorescencia en el estudio de células vivas, tejidos y otros organismos. La excitación de un fluorocromo provoca el acrecentamiento de un electrón de un estado fundamental a un estado de energía singlete excitado. Durante la relajación de la energía vibratoria a partir del estado excitado, existe la probabilidad de que se produzca un cruce entre los sistemas hasta un estado de triplete en lugar de la típica disminución de retorno al estado fundamental de singlete. Los estados de triplete son extremadamente reactivos y de vida relativamente larga, lo que permite que los fluorocromos reaccionen bajo esta condición con las células vivas o sufran degeneración molecular o reordenamiento a una especie no fluorescente. Además, los fluorocromos excitados en un estado de triplete pueden generar oxígeno en estado singlete, que reaccionará frente a una amplia variedad de grupos funcionales en biomoléculas vecinas. La luz de excitación debe penetrar la muestra a través de todos los planos focales en la trayectoria hacia el punto focal, y la mayor parte de esta luz continuará propagándose a lo largo de una distancia considerable más allá de la región focal. Por lo tanto, una población de fluorocromos excitados a lo largo de la trayectoria del haz, como es el caso con la microscopía confocal y de campo amplio, experimentará una cantidad considerable de fotoblanqueo y producirá daño celular y tisular que puede evitarse con la técnica multifotónica.
A pesar de que no se conocen muy bien los mecanismos exactos del daño celular inducido por la exposición a la luz, se ha reconocido que la disminución del fotodaño extiende radicalmente la viabilidad de las muestras biológicas investigadas con la microscopía de fluorescencia. La exposición exclusiva a la luz visible e infrarroja cercana de longitud de onda larga no parece afectar la viabilidad celular; por tanto, es probable que la mayoría del daño asociado a la microscopía multifotónica surja de la excitación, y se limite al plano focal.
Detectores para la microscopía multifotónica
En la microscopía multifotónica, los fotones emitidos a través de la fluorescencia secundaria se originan casi de forma exclusiva en el plano focal del objetivo, lo que elimina el requisito de la detección desescaneada y permite geometrías de detección más flexibles. Esta versatilidad incrementada puede derivar en una considerable optimización de la eficiencia en la detección de fluorescencia con respecto a la microscopía confocal. En un sistema con detección desescaneada, la luz recolectada por el objetivo se ve reflejada a partir de la superficie de una serie de espejos de escaneo antes de pasar a través de un estenopo hacia el detector. A medida que la resolución de la imagen aumenta, el estenopo confocal produce una gran disminución en la eficiencia de la detección, y requiere una exposición más prolongada de la muestra a la iluminación incidente, lo que aumenta la probabilidad de fotodaño y fotoblanqueo.
Con el fin de maximizar la eficiencia de la detección, por lo general los detectores no desescaneados se ubican cerca de la lente del objetivo, y el diámetro de la trayectoria de la luz debe ser mayor para detectar de manera efectiva la señal de dispersión de fluorescencia desde el interior de la muestra. Un detector común para la microscopía multifotónica es el tubo fotomultiplicador (PMT). Si se cuenta con tubos fotomultiplicadores refrigerados de fosfato-arseniuro de galio (GaAsP PMTs), es posible adquirir imágenes con una óptima relación señal-ruido, incluso con una ligera fluorescencia.

Figura 6.
Resolución en la microscopía multifotónica
La resolución en la microscopía multifotónica no supera la lograda con la microscopía confocal y, de hecho, el uso de longitudes de onda más largas (rojo a infrarrojo cercano; de 700 a 1200 nanómetros) da como resultado una función de dispersión de puntos más grande para la excitación multifotónica. Esto se traduce en una ligera reducción de la resolución tanto lateral como axial. Por ejemplo, con una longitud de onda de excitación de 700 nanómetros y un objetivo de apertura numérica de 1.3, la resolución lateral observada es de aproximadamente 0,2 micrómetros con una resolución axial correspondiente a 0,6 micrómetros. Al combinarse con la dimensión del desplazamiento Stoke, estos valores pueden variar hasta un 30 % más frente a la resolución observada con la microscopía confocal convencional en condiciones idénticas. En la práctica, la resolución confocal puede verse degradada por la apertura finita del estenopo, la aberración cromática y la alineación imperfecta del sistema óptico; todo ello sirve para reducir las diferencias de resolución entre la microscopía confocal y multifotónica. A partir de estas notas, se evidencia que las estructuras no resultas adecuadamente con un microscopio confocal, no serán mejor —o, incluso, serán peor— al ser capturadas con excitación multifotónica.
Al recopilar imágenes digitales o activar el recuento de fotones con una resolución espacial tridimensional, es fundamental hacer la distinción entre la emisión de fluorescencia que se produce dentro del volumen focal y la que se origina en el fondo. La diferenciación entre las dos señales puede lograse instrumentalmente (con instrumentación confocal o multifotónica) o por la deconvolución de un conjunto de datos tridimensionales. La capacidad para distinguir entre la emisión de fluorescencia proveniente del plano focal y la fluorescencia de fondo se rige por la relación entre señal y fondo (S/B), donde S es la cantidad o la intensidad de los fotones recogidos desde el plano focal y B representa los fotones que se originan en el fondo (planos fuera de foco). En la microscopía de escaneo confocal, se generan óptimas relaciones S/B mediante el rechazo de la señal de fondo por el estenopo confocal. Sin embargo, en la excitación multifotónica, las relaciones S/B son inherentemente amplias porque hay muy poca excitación fuera del plano focal. Los cálculos de resolución entre las técnicas multifotónicas y confocales pueden ser comparados al considerar un estenopo infinitamente pequeño cuando se llevan a cabo los cálculos confocales. En el caso de ambas técnicas, la relación entre señal y fondo habitualmente es de varios órdenes de magnitud superior a la microscopía de fluorescencia clásica de campo amplio.
Otro punto para considerar es que la excitación multifotónica permite usar fluorocromos con transiciones de absorción en la región ultravioleta de baja longitud de onda. Dado que la microscopía confocal presenta una capacidad limitada para excitar fluorocromos inferior a unos 340 nanómetros, los investigadores tienden a usar sondas dotadas de longitudes de onda mucho más largas, con resoluciones correspondientemente más bajas. En situaciones críticas, la resolución en la microscopía multifotónica puede ser mejorada al restringir las longitudes de onda del procesamiento de imágenes a través de un estenopo confocal, y también mediante el uso de un sistema de detección con resolución espacial, como una matriz de fotodiodos CCD colocada en un plano de imagen escaneada.
Características de excitación de los fluorocromos
Los fluorocromos empleados en los experimentos multifotónicos deben someterse al mismo escrutinio que los destinados a investigaciones monofotónicas. Las sondas deben dotarse de grandes secciones transversales de absorción con longitudes de onda convenientes, altos rendimientos cuánticos, una baja tasa de fotoblanqueo y el grado más ínfimo de toxicidad química y fotoquímica. Los fluorocromos también deberían ser capaces de resistir la iluminación de alta intensidad proveniente de la fuente láser sin sufrir una degradación significativa. En la mayoría de los casos, los investigadores han usado los mismos fluorocromos comunes para experimentos de dos fotones, los cuales son ampliamente aplicados como marcadores en la microscopía de fluorescencia confocal y de campo amplio.
El espectro de excitación de los fluorocromos comunes es una función del modo de excitación y la longitud de onda de los fotones incidentes. Debido a esta dependencia, el espectro de absorción de dos fotones puede (y a menudo lo hace) diferir claramente del correspondiente espectro unifotónico. Experimentalmente, la mayoría de los fluorocromos que han sido examinados son capaces de absorber la excitación bifotónica en el doble de la longitud de onda de su absorción máxima unifotónica. A pesar de ello, no existe una base fundamental para prever cuantitativamente el espectro de excitación bifotónica de un fluorocromo complejo con tan sólo examinar la sección transversal de un solo fotón. A menudo existen diferencias significativas entre los espectros de excitación unifotónica y bifotónica dedicados a las moléculas no simétricas altamente conjugadas, que a menudo se benefician de la espectroscopia molecular para proporcionar información sobre la estructura de los estados excitados. Un buen ejemplo son los derivados de los aminoácidos aromáticos de tirosina y fenilalanina, cuyas complejas secciones transversales bifotónicas son bastante diferentes de las que se muestran en la excitación unifotónica. En cambio, el espectro de dos fotones para el triptófano (Figura 2) es muy similar al perfil proyectado de la excitación unifotónica.
En el caso de los experimentos cuantitativos de reconstrucción y desconvolución tridimensionales, es necesario mediar los espectros de absorción bifotónica de los fluorocromos para asegurar que las longitudes de onda de excitación estén centradas cerca de los picos en las bandas de absorción. Si bien es posible calcular las secciones transversales de dos fotones, el proceso es complejo, y eso en el mejor de los casos. La medición experimental directa de los espectros de absorción es el método preferido; sin embargo, estos experimentos son difíciles debido a la pequeña cantidad de energía incidente absorbida contra las fluctuaciones de intensidad en la fuente de luz. Se han empleado técnicas de lentes térmicas y acústico-ópticas para determinar las secciones transversales de absorción, pero quizá un método más simple sea examinar la emisión de fotones de fluorocromos en función de un rendimiento cuántico conocido. En el diseño de nuevos experimentos bifotónicos, es necesario examinar una gama de fluorocromos que tengan picos de absorción cercanos a la mitad del valor de la longitud de onda de excitación prevista.
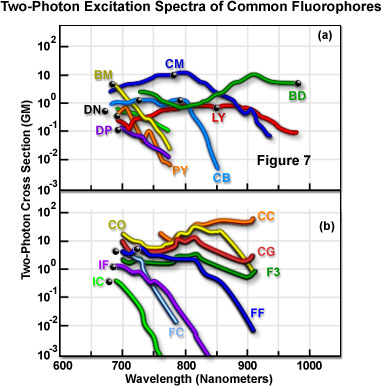
La Figura 7 presenta las características de los espectros de excitación de fluorescencia bifotónica medidos para varios fluorocromos comunes. Los datos de la Figura 7 representan secciones transversales de acción bifotónica, derivadas del producto de la eficiencia cuántica de la emisión de fluorescencia y la sección transversal de absorción bifotónica. Los espectros se registraron usando la luz polarizada emitida de forma lineal por un láser con bloqueo de modo de titanio-zafiro. En cada espectro, el punto negro representa el doble de la longitud de onda del nivel máximo de absorción de un solo fotón del fluorocromo. La Tabla 1 es clave para obtener los códigos de los nombres de dos letras que se presentan junto a cada espectro en la Figura 7. Las curvas representan las secciones transversales espectrales de la excitación bifotónica del fluorocromo.
Espectros de excitación de fluorescencia bifotónica de fluorocromos
| Nombre del fluorocromo (abreviatura) | Longitud de onda de excitación (nanómetros) |
|---|---|
| [BM] Benceno, p-bis (o-metilestiril) | 691 |
| [CB] Sal trisódica de hidrácida Cascade Blue | 750 |
| [YL] Sal de amonio CH Lucifer Yellow | 860 |
| [BD - BODIPY] Sal disódica del ácido 4-4-diflouro-1,3,5,7,8-pentametil-4-bora-3a, 4a-diaza-s-indacenos-2,6-disulfónico | 920 |
| [DP - DAPI no unido al ADN] Diclorhidrato de 4',6-diamidino-2-fenilindol | 700 |
| [DN - DANSYL] 5-dimetilaminonaftaleno-1-sulfonilhidrazina | 700 |
| [PY] 1,2-bis-(1-pirenodecanoil)-sn-glicero-3-fosfocolina | 700 |
| [CM] Cumarina 307 | 776 |
| [IC] Indo-1 con Ca++ | 700 |
| [IF] Indo-1 sin Ca++ | 700 |
| [FC] Fura-2 con Ca++ | 700 |
| [FF] Fura-2 sin Ca++ | 720 |
| [CG] Calcio verde-1 con Ca++ | 725 |
| [CO] Calcio naranja con Ca++ | 800 |
| [CC] Calcio rosado con Ca++ | 850 |
| [F3] Fluo-3 con Ca++ | 800 |
Tabla 1
Las mediciones transversales indican una tendencia en la que el pico de excitación dedicado a la absorción bifotónica es muy similar o se desplaza hacia el azul con respecto al perfil de un solo fotón (Figura 7). Las longitudes de onda promedio más cortas pueden ser mejor para acoplar la excitación del fluorocromo al rango de longitud de onda disponible en los láseres pulsados de bloqueo de modo. Otro aspecto coherente de los espectros de absorción bifotónica es que, por lo general, son mucho más amplios que sus contrapartes de un solo fotón. Esto mitiga las limitaciones experimentales si se aumenta el rango de longitudes de onda adecuadas para la excitación y si se mejora la capacidad de excitar simultáneamente dos fluorocromos dotados de secciones transversales bifotónicas superpuestas, pero con espectros unifotónicos muy separados. Las mediciones de las secciones transversales trifotónicas indican que, en general, son muy similares a los correspondientes espectros de un solo fotón.
Si bien los espectros de absorción a menudo difieren en la excitación de uno y dos fotones, no parece que esto influye en las otras propiedades de fluorescencia, como la vida útil, las longitudes de onda de emisión y el índice de cruce entre sistemas. Dicha similitud indica que se alcanzan los mismos estados excitados por fluorescencia mediante la absorción lineal o no lineal, y que, una vez excitado el fluorocromo, éste se comportará de igual forma sin considerar el modo de excitación. Estos ocupantes también son válidos para la excitación trifotónica, lo que permite a los investigadores usar métodos radiométricos y espectroscópicos bien establecidos en la mayoría de los experimentos multifotónicos.
Daño térmico y fotográfico en la excitación multifotónica
Todas las formas de microscopía de fluorescencia sufren fotodaño en las células vivas, cuyo grado depende de la longitud de onda de excitación, la duración de la exposición y la naturaleza química de los fluorocromos usados como sondas celulares. El daño inducido por la iluminación de excitación puede categorizarse en dos grupos: daño término y degradación por reacciones químicas. Los efectos secundarios fotoquímicos causados por las reacciones bioquímicas, como resultado de la excitación del fluorocromo, no son bien comprendidos y varían ampliamente entre los tipos de células y tejidos. Por otro lado, el daño térmico surge principalmente a partir de dos mecanismos que ocurren debido a la absorción de un solo fotón por el agua y por la absorción de dos fotones de los fluorocromos en la región focal.
En la mayoría de las células estudiadas (particularmente las células de mamíferos), casi no hay absorción de la radiación de excitación en el infrarrojo cercano de longitud de onda larga por los fluorocromos intrínsecos usados en la fluorescencia multifotónica. Sin embargo, el líquido intracelular e intercelular que rodea las células y los tejidos puede absorber cantidades significativas de iluminación infrarroja e infrarroja cercana, lo que produce un exceso de calor que puede dañar la viabilidad de las muestras biológicas. Por otro lado, cuando el entorno biológico acuoso se ilumina con las longitudes de onda ultravioleta y visible más cortas, usadas en la microscopía de fluorescencia confocal y de campo amplio, el líquido circundante no absorbe una cantidad significativa de calor.
El calentamiento debido a la absorción de un solo fotón por el agua se produce a lo largo de la trayectoria del haz, tanto por encima como por debajo del plano focal. En condiciones multifotónicas promedio controladas, se ha calculado que el aumento inducido de la temperatura oscila entre 0,065 y 1,1 grados centígrados en 700 y 1000 nanómetros, respectivamente. Estos cálculos son conforme con las mediciones térmicas ejecutadas en 1064 nanómetros mediante la excitación láser de pinzas ópticas. En situaciones en las que el haz de luz excitante se mantiene estacionario, puede producirse un mayor calentamiento, lo que edifica la relación logarítmica con el tiempo. El calentamiento debido a la absorción de fluorocromos se localiza bastante en la región focal entre los experimentos de excitación multifotónica. La posterior liberación del calor ocurre uniformemente dentro de una región esféricamente simétrica que rodea el volumen focal y no produce una cantidad significativa de calor, incluso con altas concentraciones de fluorocromos.
Conclusiones
La microscopía de fluorescencia multifotónica está convirtiéndose en uno de los métodos de predilección para el procesamiento dinámico de imágenes provenientes de células y tejidos de animales vivos destinados a la experimentación. La técnica es particularmente útil cuando se observa la dinámica de las células ubicadas en lo profundo de la muestra. Por otra parte, los efectos secundarios en la excitación multifotónica, como el fotoblanqueo y el fotodaño, disminuyen y ocurren solo en la región inmediata entorno al volumen del foco.
Si bien, la fototoxicidad en las células es un fenómeno poco conocido, este ocurre seguido en la mayoría de las formas de microscopía de fluorescencia. Por consiguiente, tanto la energía cuántica como la absorción intrínseca más bajas de las longitudes de onda más largas, que se emplean en la microscopía multifotónica, sirven para reducir los efectos nocivos de la luz en las células y tejidos vivos, lo que abre camino a las investigaciones de la dinámica celular. Un obstáculo importante para la investigación de la microscopía multifotónica es el alto costo del equipamiento, en particular con los sistemas de láser pulsado con bloqueo de modo necesarios para la excitación bifotónica y trifotónica. Los sistemas láser ultrarrápidos más populares de uso general son los láseres de titanio-zafiro. La optimización que se experimenta en la sintonización de la longitud de onda del láser pulsado de titanio-zafiro (700 a 1300 nanómetros) lo hace mucho más versátil. Esta disponibilidad fomentará la aplicación generalizada de esta técnica en todas las ciencias biológicas.
Sorry, this page is not
available in your country.



