Découverte des mécanismes de régulation qui déterminent le bon assemblage des cils motiles des cellules multiciliées à l’aide de l’imagerie de cellules vivantes à grande vitesse
Cette note d’application présente les cils cellulaires et explique la façon dont ils se forment, se déplacent et s’assemblent. Découvrez comment des travaux de recherches recourant à l’imagerie de cellules vivantes à grande vitesse révèlent les mécanismes de régulation qui déterminent le bon assemblage des cils motiles des cellules multiciliées.
Présentation des cils cellulaires
Les cils cellulaires sont des structures ressemblant à des poils qui remplissent diverses fonctions essentielles dans les cellules animales. Parmi ces fonctions, on peut citer les fonctions de locomotion, de détection de l’environnement et d’alimentation. Les cils cellulaires ont un diamètre d’environ 0,2 μm et une longueur de 10 μm.
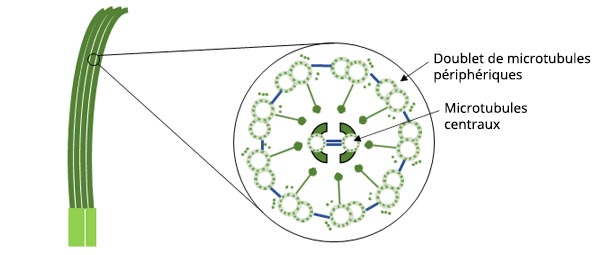
Figure 1 : Schéma de la structure des cils cellulaires
Dans la fonction de locomotion, les cils cellulaires agissent comme des pagaies qui vont déplacer la cellule. Les paramécies, des organismes microscopiques qui vivent dans les étangs et les ruisseaux, sont couvertes de cils cellulaires qui battent comme des rames miniatures pour propulser ces créatures dans l’eau.
Chez les animaux de plus grande taille, les cils cellulaires assurent des fonctions plus complexes. Ils fournissent une source de propulsion pour donner lieu à un mouvement, par exemple pour pousser le spermatozoïde et l’ovule à s’unir. Certains cils fonctionnent comme des antennes pour détecter des stimuli externes.
La plupart des cellules animales, y compris presque tous les types de cellules du corps humain, ont une surface ciliée. Les cils cellulaires se répartissent en cils motiles et en cils statiques, et la plupart des cils motiles existent en groupes de plusieurs cils. Les cellules multiciliées (CMC) sont des cellules épithéliales qui contiennent des centaines de cils motiles utilisés pour propulser le liquide à la surface de la cellule. Elles sont principalement distribuées dans les tissus épithéliaux, ces tissus minces qui tapissent diverses surfaces de l’organisme. Dans les voies respiratoires, les CMC nettoient et éliminent le mucus et d’autres irritants de l’organisme. Dans la moelle épinière et les ventricules cérébraux des adultes, elles déplacent le liquide céphalorachidien dans le système nerveux central.
Lorsque les cils cellulaires ne fonctionnent pas correctement, cela peut avoir un large éventail d’effets sur la santé humaine. Les maladies génétiques entraînant des anomalies des cils cellulaires humains sont collectivement appelées ciliopathies. La liste des ciliopathies ne cesse de croître, avec un grand nombre de maladies rares qui affectent souvent plusieurs organes du corps. Comprendre comment les cils cellulaires se forment, se déplacent et s’assemblent peut aider les chercheurs à en apprendre davantage sur ces maladies.
La relation entre les centrioles et les cils cellulaires
Les centrioles sont des structures en forme de tonneau qui sont essentielles à la formation des cils et des centrosomes. Dans les cellules, les centrioles apparaissent généralement par deux. Certaines protéines spécifiques peuvent se rassembler autour de chaque paire de centrioles pour former des centrosomes, des organites responsables de la croissance et de l’ancrage des microtubules dans les cellules. Les centrosomes agissent comme les principaux centres d’organisation des microtubules de la cellule et comme un régulateur de la division cellulaire.
Lorsque les cellules copient leurs chromosomes avant la division, les deux centrioles se divisent pour faire pousser un centriole « fils » de chaque côté et former ainsi deux centrosomes. Ensuite, ces deux centrosomes, composés d’une paire de centrioles père-fils et de microtubules radiaux autour d’eux, forment des pôles de fuseau mitotique responsables de la distribution du matériel génétique (chromosomes) de manière égale dans les deux cellules filles. Après la division, chaque cellule fille acquiert le même nombre de chromosomes et un centrosome.
Lorsque les cellules sont dans la phase de non-prolifération, le centriole père est ancré à la membrane cellulaire, et les cils se développent à son extrémité. Lorsque la cellule est sur le point de proliférer, les cils raccourcissent progressivement et disparaissent complètement avant la division cellulaire. Cela permet de libérer les centrosomes dans le cytoplasme pour former des fuseaux mitotiques. De cette façon, le centriole joue deux rôles différents pour former des centrosomes et des cils cellulaires.
Comprendre pourquoi plusieurs nouveaux centrioles apparaissent spontanément
Étant donné que les centrioles se dupliquent une seule fois par cycle cellulaire, d’où vient le grand nombre de centrioles rencontré dans les CMC (comme les cils des paramécies) ? Dès les années 1960, les biologistes cellulaires ont pu observer par microscopie électronique que le nombre de centrioles augmentait rapidement pendant la formation des CMC. Ils ont également constaté de façon inattendue qu’un plus grand nombre de centrioles se formait autour de nombreuses structures circulaires appelées deutérosomes.
Plusieurs dizaines d’années plus tard, on ne savait toujours pas pourquoi de nouveaux centrioles apparaissaient spontanément autour des deutérosomes. Ce n’est que suite à une étude menée dans notre laboratoire en 2013 que le mystère a enfin pu être percé1. Nous avons constaté qu’une paire de protéines homologues, Deup1 et Cep63, présentes chez les animaux supérieurs régulait les deux modes d’amplification des centrioles au cours de la formation de novo (c.-à-d., respectivement, l’assemblage de centrioles sans centrioles préexistants et la formation dépendante d’un centriole père dans le processus de production de cils multiples). Cela a clarifié l’origine des nombreux nouveaux centrioles dans les CMC.
L’assemblage précis des cils motiles
Les cils motiles contiennent des centaines de constituants protéiques synthétisés en grandes quantités lors de la différenciation des cellules multiciliées. Que leur assemblage se fasse de façon spontanée ou qu’il soit organisé de manière spécifique et stocké à la demande est une autre question scientifique importante et restée sans réponse.
Les résultats que nous avons publiés révèlent que les matériaux fibrogranulaires (MFG), une structure subcellulaire unique aux CMC, jouent un rôle important en tant qu’organisateurs dans la régulation précise de l’assemblage des cils motiles2.
Les granules fibreux sont des structures subcellulaires qui ne sont présentes que dans les CMC des animaux supérieurs. Bien qu’ils aient été découverts dans des études de microscopie électronique menées dans les années 1960, leur composition et leur fonction sont encore peu claires, mis à part que l’on sait qu’ils contiennent la protéine de la matrice péricentrosomique (PCM1). En combinant la protéomique, la microscopie à super-résolution, la microscopie de cellules vivantes à grande vitesse et diverses techniques de microscopie électronique, nous avons fait les observations suivantes :
- Les granules fibreux sont de grands organites sans membrane aux caractéristiques de liquide formés par une séparation de phase de PCM1, qui adhèrent aux deutérosomes et enrichissent leur structure en forme de mûre avec de nombreuses protéines structurales de stroma et de cils spécifiques. Ces protéines structurales sont localisées dans la matrice et les cils à différents stades de la ciliogenèse.
- Une réduction génétique artificielle de la concentration en protéine PCM1 empêche les CMC de former des particules fibreuses et conduit à une altération du nombre, de la taille et de la distribution des deutérosomes. Bien que ces cellules aient finalement formé le même nombre de cils que les cellules témoins, la structure de leurs cils et de leur matrice était significativement défectueuse. Cela a entraîné de graves anomalies dans la façon dont les cils s’agitent ou de leur capacité à s’agiter.
- Une analyse approfondie a révélé que les composants enrichis dans les particules fibreuses étaient localisés à l’avance dans la matrice et que la structure de la base des microtubules centraux des cils était significativement étendue. Cela indique que l’ordre chronologique de leur assemblage a été rompu, ce qui a affecté la précision et la fonction de la structure des cils motiles.
Par conséquent, les granules fibreux remplissent la fonction d’organisateur pendant la différenciation des CMC, à la fois comme salle de stockage pour les composants liés aux cils et pour l’adhérence au deutérosome et l’enrichissement près de la base des nouveaux cils pour assurer le bon assemblage de la structure fine des cils motiles.
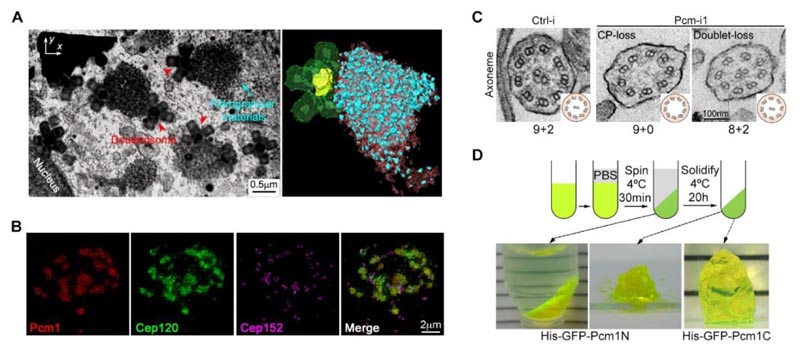
Figure 2. (A) La microscopie électronique à balayage et à faisceau d’ions focalisé (FIB-SEM) a révélé la distribution spatiale des particules fibreuses et des deutérosomes dans les cellules multiciliées. (B) La structure des particules fibreuses peut enrichir la matrice en protéine structurale Cep120. (C) Le fait de réduire la concentration en protéine PCM1 entraîne des défauts évidents dans l’ultrastructure des cils mobiles. (D) Des fragments de PCM1 ont une propriété de séparation de phase2.
Analyse du bon assemblage des cils motiles de cellules multiciliées
Dans nos recherches2, nous avons utilisé diverses technologies d’imagerie, y compris la microscopie à super-résolution, la microscopie de cellules vivantes à grande vitesse et la microscopie électronique. Le microscope à super-résolution IXplore™ SpinSR d’Evident a joué un rôle primordial dans ce travail. Vous trouverez ci-dessous une présentation détaillée de l’application pratique du système IXplore SpinSR dans nos recherches.
Nous avons réalisé une imagerie de cellules vivantes à deux canaux sur des cellules épithéliales épendymiques ventriculaires de souris (mEPC) infectées par un adénovirus et un lentivirus pour exprimer les protéines de fusion SNAP-Deup1 et GFP-PCM1 sous le contrôle du promoteur de Deup1.
Nous avons observé le comportement dynamique de la protéine GFP-PCM1 dans les mEPC en utilisant l’imagerie en temps réel in vivo sur le système IXplore SpinSR. Comme l’expression de la protéine SNAP-Deup1 est contrôlée par le promoteur de Deup1, l’apparition de points SNAP-Deup1 positifs indique que la cellule subit une expansion des centrioles.
Les résultats d’imagerie montrent :
- Une colocalisation claire entre les deutérosomes marqués par SNAP-Deup1 et les MFG marqués par GFP-PCM1 dans les mEPC. Ils sont de tailles différentes mais se déplacent ensemble.
- Les plus gros points de MFG semblent envelopper complètement le deutérosome, et parfois plusieurs deutérosomes sont confinés à un gros point de MFG.
- Lorsque deux points de MFG se rencontrent, ils fusionnent graduellement en un point de MFG plus gros, dans lequel leurs deutérosomes internes sont fusionnés (figure 3). Par conséquent, ces expériences démontrent que les MFG ont des propriétés liquides et se lient étroitement aux deutérosomes.
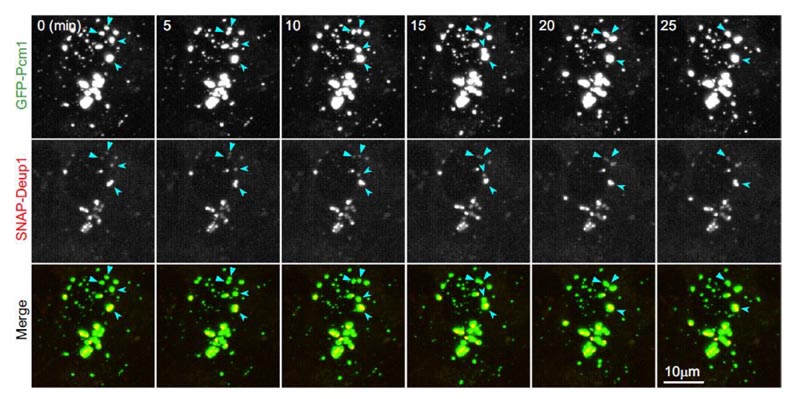
Figure 3. Les MFG des mEPC vivantes ont des propriétés de liquide et se lient étroitement au deutérosome. Le point de MFG a des propriétés de liquide et se lie au deutérosome. Des images des mEPC vivantes sont prises en temps réel toutes les cinq minutes le jour 3. GFP-PCM1 et SNAP-Deup1 indiquent respectivement les MFG et les deutérosomes. Les flèches indiquent l’événement de fusion des deux MFG dans la séquence d’images2 .
Conditions d’imagerie
Système : système IXplore SpinSR (mode confocal)
Objectif : APON60XOTIRF / ON 1,49
Temps d’exposition : 100 millisecondes
Durée : toutes les cinq minutes pendant 65 minutes
De plus, nous avons incubé des cellules épithéliales trachéales de souris (CETS) cultivées dans des plaques Transwell avec 200 nM de SIR-tubuline pour marquer leurs cils multiples par fluorescence. En utilisant l’imagerie à grande vitesse sur le système IXplore SpinSR pour capturer les trajectoires motrices des CETS GFP-Centrin1 positives, nous avons constaté que les cils motiles présentaient principalement une oscillation régulière ondulatoire dans les cellules témoins, tandis que les cils motiles tournaient de façon irrégulière ou perdaient complètement leur capacité à bouger dans les CETS à concentration réduite en PCM1 (figure 4).

Related Videos
Figure 4. La suppression de PCM1 entraîne une motilité ciliaire anormale des CETS2 .
Conditions d’imagerie
Système : système IXplore SpinSR (mode super-résolution)
Objectif : APON60XOTIRF (ON 1,49)
Temps d’exposition : 14 millisecondes
Durée d’acquisition : intervalle de 15 millisecondes, 3 secondes
Comment le système de microscopie à super-résolution IXplore SpinSR a facilité l’expérience
Le système d’imagerie à super-résolution IXplore SpinSR permet l’observation en temps réel des échantillons vivants. Il utilise la technologie confocale à disque rotatif pour une acquisition d’images rapide avec une phototoxicité minimale pour les cellules vivantes.
L’observation en imagerie à intervalles est un outil couramment employé en recherche biologique pour relever les changements qui s’opèrent dans les structures ainsi que les phénomènes intracellulaires, mais elle impose des paramètres adaptés aux conditions d’observation nécessaires pour rendre compte de ces changements. Le système IXplore SpinSR permet l’imagerie des cellules vivantes en super-résolution pour capter les changements rapides et les phénomènes recherchés en minimisant autant que possible la phototoxicité.
En plus des performances de l’équipement, le traitement des données à grande vitesse du système IXplore SpinSR permet d’obtenir des images en super-résolution dans une fenêtre d’affichage en direct.
Principales caractéristiques :
- Résolution horizontale : 110 nm (combinaison de disques Sora)
- Balayage à grande vitesse
- Imagerie à super-résolution multicolore
- Configuration facile
| RemerciementsCet article a été coécrit avec Yan Xiumin du Shanghai Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences. |

Références bibliographiques
1. Huijie Zhao et al. « The Cep63 paralogue Deup1 enables massive de novo centriole biogenesis for vertebrate multiciliogenesis », Nature Cell Biology, no 15 (Déc. 2013): 1434–1444. doi: 10.1038/ncb2880.
2. Huijie Zhao et al. « Fibrogranular materials function as organizers to ensure the fidelity of multiciliary assembly », Nature Communications, n° 12: 1273 (Fév. 2021). doi.org/10.1038/s41467-021-21506-8.
Produits utilisés pour cette application
Sorry, this page is not
available in your country.