Revealing the Regulatory Mechanisms for the Proper Assembly of Motile Multicilia Using High-Speed Live-Cell Imaging
This application note introduces cilia and explores how they form, move, and assemble. Learn how research using high-speed live cell imaging is revealing the regulatory mechanisms for the proper assembly of motile multicilia.
Introduction to Cilia
Cilia are hair-like structures that serve a wide variety of essential functions in animal cells. These functions include locomotion, sensing the environment, and feeding. Cilia are about 0.2 μm in diameter and 10 μm long.
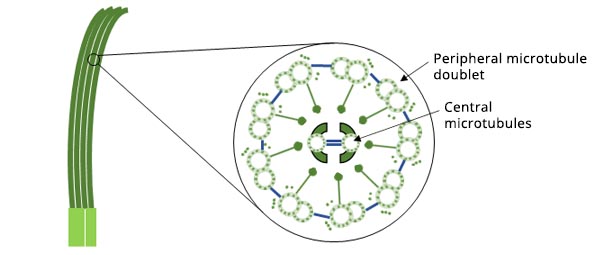
Figure 1. Schematic diagram of the structure of cilia.
In locomotion, cilia serve as paddles that move the cell. Paramecia, microscopic organisms that live in ponds and streams, are covered with cilia that beat like miniature oars to propel the creatures through water.
In larger animals, cilia take on more complex functions. They provide a source of power for movement, such as pushing sperm and egg to union. Some cilia work like antennas to feel external stimuli.
Most animal cells, including almost all types of cells in the human body, have cilia on the surface. Cilia are divided into motile cilia and static cilia, and most motile cilia exist in clusters. Multiciliated cells (MCCs) are epithelial cells that contain hundreds of motile cilia used to propel fluid over the surface of the cell. They are mainly distributed in epithelial tissues—thin tissues that cover various surfaces and linings throughout the body. In the respiratory tract, MCCs clean and clear mucus and other irritants from the body. In the spinal cord and brain ventricles of adults, they move cerebrospinal fluid through the central nervous system.
When cilia fail to work properly, they can cause a wide range of effects on human health. Genetic diseases causing human cilia abnormalities are collectively referred to as ciliopathies. The list of ciliopathies continues to grow with a range of rare diseases that often affect multiple organs within the body. Understanding more about how cilia form, move, and assemble can help researchers learn more about these diseases.
The Relationship between Centrioles and Cilia
Centrioles are barrel-shaped structures that are essential for the formation of both cilia and centrosomes. In cells, centrioles usually appear in pairs. Some specific proteins can gather around each pair of centrioles to form centrosomes, organelles that are responsible for growing and anchoring microtubules in cells. Centrosomes act as the main microtubule-organizing centers of the cell and regulator of cell division.
As cells copy their chromosomes before division, the two centrioles split to grow a daughter centriole on each side to form two centrosomes. Then these two centrosomes, composed of a mother-daughter centriole pair and the radial microtubules around them, form spindle poles that are responsible for distributing genetic material (chromosomes) equally into the two daughter cells. After division, each daughter cell acquires the same number of chromosomes and a centrosome.
When cells are in the nonproliferation phase, the mother centriole is anchored to the cell membrane, and cilia grow at the tip of it. When the cell is about to proliferate, the cilia will gradually shorten and disappear completely before cell division. This lets the centrosomes be released into the cytoplasm to form spindles. In this way, the centriole plays two different roles to form centrosomes and cilia.
Understanding Why Multiple New Centrioles Appear Spontaneously
Since centrioles duplicate once per cell cycle, where do the large number of centrioles in MCCs (such as cilia on Paramecia) come from? As early as the 1960s, cell biologists used electron microscopy to observe that centrioles would expand rapidly during the formation of MCCs. They also unexpectedly found that more centrioles formed around many circular structures called deuterosomes.
Decades later, little is known about why new centrioles appear spontaneously around deuterosomes. It was not until a study in our lab in 2013 that the mystery was uncovered.1 We found that a pair of homologous proteins, Deup1 and Cep63, in higher animals regulated the two centriole amplification modes of de novo formation (i.e., assembly of centrioles without any preexisting centrioles) and mother-centriole dependent formation in the process of producing multiple cilia, respectively. This clarified the origin of many new centrioles in MCCs.
The Precise Assembly of Motile Cilia
Motile cilia contain hundreds of protein components that are synthesized in large quantities during multiciliated cell differentiation. Whether they are a spontaneous assembly or organized in a specific way and stored on demand is another important and unanswered scientific question.
Our published findings reveal that fibrogranular materials (FGMs), a subcellular structure unique to MCCs, play an important role as organizers in regulating the precise assembly of motile cilia.2
Fibrous granules are subcellular structures that occur only in higher animal MCCs. Although they have been discovered in electron microscopy studies in the 1960s, their composition and function are still unclear except for the knowledge that they contain the pericentrosome matrix protein (PCM1). Combining proteomics, super-resolution microscopy, live-cell high-speed microscopy, and multiple electron microscopy techniques, we found:
- Fibrous granules are large, liquid-characterized, membraneless organelles, formed by PCM1 phase separation, that adhere to deuterosomes and enrich their mulberry-like structure with many structural proteins of specific stroma and cilia. These structural proteins are localized into the matrix and cilia at different stages of ciliogenesis.
- Artificially knocking down the protein level of PCM1 prevents MCCs from forming fibrous particles and leads to an abnormal deuterosome number, size, and distribution. Although these cells eventually formed the same number of cilia as control cells, the structure of their cilia and matrix was significantly defective. This resulted in severe abnormalities in the way cilia wiggle and their ability to wiggle.
- In-depth analysis revealed that the components enriched in the fibrous particles were localized to the matrix in advance, and the structure of the central microtubule base of the cilia was significantly extended. This indicated that the chronological order of their assembly was broken, affecting the accuracy and function of the motile cilia structure.
Therefore, fibrous granules perform the function of an organizer during the differentiation of MCCs, both as a storage room for cilia-related components and to adhere to the deuterosome and enrich near the base of the new cilia to ensure the proper assembly of the fine structure of motile cilia.
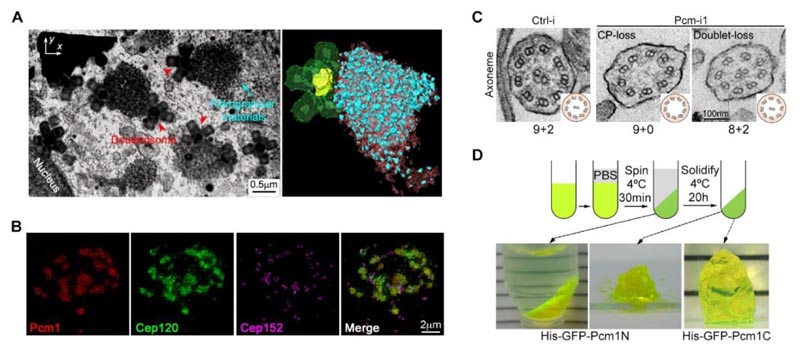
Figure 2. (A) Focused ion beam-scanning electron microscopy (FIB-SEM) showed the spatial distribution of fibrous particles and deuterosomes in multiciliated cells. (B) Fibrous particulate structure can enrich the structural protein Cep120 of the matrix. (C) Knocking down the protein level of PCM1 leads to obvious defects in the ultrastructure of the moving cilia. (D) PCM1 fragments have the property of phase separation.2
Analyzing the Proper Assembly of Motile Multicilia
In our research,2 we used a range of imaging technologies, including super-resolution microscopy, live-cell high-speed microscopy, and electron microscopy. Central to this work was the IXplore™ SpinSR super-resolution microscope from Evident. The following is a detailed introduction to the practical application of the IXplore SpinSR system in our research.
We performed two-channel live-cell imaging of mouse ventricular ependymal epithelial cells (mEPCs) infected with adenovirus and lentivirus virus to express SNAP-Deup1 under GFP-PCM1 and Deup1 promoters.
We observed the dynamic behavior of GFP-PCM1 in mEPCs using in vivo live imaging on the IXplore SpinSR system. Since the expression of SNAP-Deup1 is controlled by the Deup1 promoter, the appearance of SNAP-Deup1-positive dots indicates that the cell is undergoing centriole expansion.
The imaging results showed that:
- There was clear colocalization between SNAP-Deup1-labeled deuterosomes and GFP-PCM1-labeled FGMs in mEPCs. They are different sizes but move together.
- The larger size of the FGM points seems to completely wrap the deuterosome, and sometimes several deuterosomes are confined to a large FGM point.
- When two FGM points meet, they gradually merge into a larger FGMs point, where their internal deuterosomes are fused together (Figure 3). Therefore, these experiments have proved that FGMs have liquid properties and tightly bind to deuterosomes.
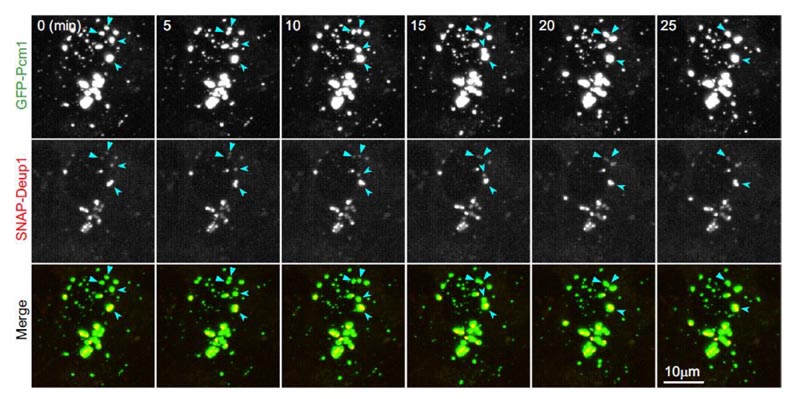
Figure 3. FGMs in living mEPCs have liquid properties and bind tightly to the deuterosome. The FGMs point has liquid properties and binds to the deuterosome. Live mEPCs are imaged in real time every five minutes on Day 3. GFP-PCM1 and SNAP-Deup1 indicate FGMs and deuterosomes, respectively. The arrows indicate the fusion event of the two FGMs in the image sequence.2
Imaging Conditions
System: IXplore SpinSR system (confocal mode)
Objective lens: APON60XOTIRF/ 1.49 NA
Exposure time: 100 milliseconds
Duration: Every five minutes for 65 minutes
In addition, we incubated mouse tracheal epithelial cells (MTECs) grown in Transwells with 200 nM SiR-tubulin to fluorescently label their multicilia. Using high-speed imaging on the IXplore SpinSR system to capture the motor trajectories of GFP-Centrin1-positive MTECs, we found that motile cilia mainly showed a regular wave-like swing in control cells, while motile cilia rotated irregularly or completely lost their ability to move in PCM1-knocked MTECs (Figure 4).
Related Videos

Figure 4. PCM1 deletion leads to abnormal ciliary motility of MTECs.2
Imaging Conditions
System: IXplore SpinSR system (super-resolution mode)
Objective lens: APON60XOTIRF (1.49 NA)
Exposure time: 14 milliseconds
Acquisition time: 15 milliseconds interval, 3 seconds
How the IXplore SpinSR Super-Resolution Microscope System Facilitated the Experiment
The IXplore SpinSR super-resolution imaging system enables live observation of specimens. It employs spinning disk confocal technology for fast image acquisition with minimal phototoxicity to living cells.
Time-lapse observation is a common tool in biological research to capture changes in intracellular structures and phenomena, but it requires settings tailored to the observation conditions needed to capture those changes. The IXplore SpinSR system enables live-cell super-resolution imaging to capture fast changes and capture the correct phenomena by minimizing phototoxicity as much as possible.
In addition to hardware performance, the IXplore SpinSR system's high-speed data processing enables super-resolution images in a live display window.
Key features:
- Horizontal resolution: 110 nm (Sora disk combination)
- High-speed scanning
- Multicolor super-resolution imaging
- Easy setup
| AcknowledgementsThis article was cowritten with Yan Xiumin, Shanghai Institute of Biochemical Cell Biology, Chinese Academy of Sciences. |

References
1. Huijie Zhao et al. “The Cep63 paralogue Deup1 enables massive de novo centriole biogenesis for vertebrate multiciliogenesis.” Nature Cell Biology, no. 15 (Dec. 2013): 1434–1444. doi: 10.1038/ncb2880.
2. Huijie Zhao et. al. “Fibrogranular materials function as organizers to ensure the fidelity of multiciliary assembly.” Nature Communications, no. 12: 1273 (Feb. 2021). doi.org/10.1038/s41467-021-21506-8.
Products related to this application
Sorry, this page is not
available in your country.